摘要:本研究从传统发酵食品中筛选产脂肪酶乳酸菌,结合干酪浆快速成熟模型得到产内酯化合物性能较好的乳酸菌,将其作为附属发酵剂应用于切达干酪的制备,采用感官评价、气相色谱-质谱(ga
本研究从传统发酵食品中筛选产脂肪酶乳酸菌,结合干酪浆快速成熟模型得到产内酯化合物性能较好的乳酸菌,将其作为附属发酵剂应用于切达干酪的制备,采用感官评价、气相色谱-质谱(gas chromatography-mass spectrometry, GC-MS)联用技术比较添加产脂肪酶乳酸菌的附属发酵剂和未添加附属发酵剂干酪在4、10、14℃成熟过程中(150 d)风味品质及内酯化合物含量的变化规律。结果表明,产脂肪酶乳酸片球菌4D和格氏乳球菌Y3具有较好的产内酯性能。经感官评价发现,与未添加附属发酵剂的切达干酪相比,添加乳酸片球菌4D的干酪在10、14℃成熟90 d时分别具有香气强度最高的奶香味、果香;添加格氏乳球菌Y3的干酪在14℃成熟120 d具有最高的奶香味香气强度值。利用GC-MS技术共鉴定出4种内酯类化合物,添加乳酸片球菌4D和格氏乳球菌Y3的切达干酪在3种恒温条件下成熟时,δ-癸内酯和δ-十二内酯的含量均高于未添加附属发酵剂的切达干酪,且能够促进δ-辛内酯的形成。此外,仅在格氏乳球菌Y3发酵的切达干酪中鉴定出γ-丁内酯。通过探究干酪成熟温度和时间对内酯形成的影响,发现在14℃条件下干酪的成熟效果最好,添加乳酸片球菌4D的切达干酪在90 d时总内酯含量达到最高,添加格氏乳球菌Y3的切达干酪则是在120 d时达最高。
关键词:乳酸菌;脂肪酶;切达干酪;内酯化合物;附属发酵剂
论文《高产脂肪酶乳酸菌的筛选及其对切达干酪内酯类化合物形成的影响》发表在《食品科学》,版权归《食品科学》所有。本文来自网络平台,仅供参考。
一、引言
切达干酪是以牛奶为原料,经浓缩、发酵等工序制成具有独特风味的发酵乳制品[1]。对于消费者而言,切达干酪产品的风味品质是影响是否购买干酪产品的重要因素[2]。附属发酵剂是指在干酪加工过程中除了添加主发酵剂乳酸菌外添加的一类可用于提高干酪风味品质或促进成熟的选定微生物[3]。乳酸菌因在发酵过程可产生乳酸促进凝乳便于排出乳清,还可通过碳水化合物代谢、脂肪酸代谢形成的醛、酮、内酯化合物赋予干酪奶香、果香风味而备受关注[4-5]。例如Gobbetti等[6]的研究表明,在切达干酪中附属发酵剂可促进蛋白水解改善其风味品质。康优等[7]将乳酸片球菌(Pediococeus acidilaetici)AS185制成附属发酵剂用于切达干酪的发酵成熟,与仅添加商业发酵剂干酪相比,添加附属发酵剂的干酪具有更好感官属性。
内酯类化合物在干酪中的含量较低,但其嗅觉阈值低,属于干酪的关键风味化合物[8]。牛乳中的甘油三酯在脂肪酶的作用下分解成脂肪酸,并进一步通过β-氧化及环化生成γ-或δ-内酯类化合物[9]。利用微生物所产的脂肪酶改善产品风味可满足消费者对食品天然、健康的需求,因此这种方式也备受关注[10]。如研究人员发现将米曲霉(Aspergillus oryzae)所产的脂肪酶与嗜热链球菌(Streptococcus thermophilus)、鼠李糖乳酪杆菌(Lacticaseibacillus rhamnosus)等乳酸菌混合发酵,可显著提高酸奶的风味品质[11]。洛克菲特青霉菌(Penicillium roqueforti)所产的脂肪酶在蓝纹干酪成熟过程中能够提高干酪的质量、外观与风味[12]。产脂肪酶微生物还能够在不影响产品的理化性质情况下促进干酪的成熟[13]。但不同微生物所产脂肪酶在酶学特性上存在差异[14],因此探究干酪的加工参数(如温度、成熟时间)对于利用微生物改善干酪风味十分重要。目前,国内外研究学者发现瑞士乳杆菌(Lactobacillus helveticus)、干酪乳酪杆菌(Lacticaseibacillus casei)在干酪成熟14 d时可促进其中苯甲醛、乙偶姻的形成[15]。在成熟60 d时,经副干酪乳酪杆菌、发酵乳杆菌发酵的干酪中乙酸、己酸乙酯、苯乙醇含量较高[16]。
切达干酪的成熟过程以及风味的稳定形成耗时较长,因此国内外学者通常建立干酪浆快速成熟模型以实现快速准确评价干酪成熟过程中的风味品质变化[17-18]。干酪浆体系能够加速干酪的成熟,如Boran等[19]利用干酪浆加速了干酪的成熟,并实现干酪风味的改善。这一加速过程是由于干酪浆具有较高的含水量和成熟温度(30℃),从而加快体系内微生物的生化代谢[20]。干酪浆快速成熟模型也可用于评估乳酸菌在干酪发酵过程的作用,如李春燕等[21]利用干酪浆快速成熟模型筛选出具有较好蛋白水解能力的附属发酵剂乳酸菌。
由于缺乏优质的产脂肪酶乳酸菌资源及其对切达干酪内酯化合物贡献尚不明晰,因此有必要筛选高产脂肪酶乳酸菌,开发相应的附属产香发酵剂,并探究工艺参数对干酪风味调控的影响。基于此,本研究利用三丁酸甘油酯透明圈法结合铜皂法从不同发酵样品中筛选高产脂肪酶乳酸菌,对其生长特性进行分析。进一步制作干酪浆快速成熟模型探究不同高产脂肪酶乳酸菌的产内酯类化合物性能,筛选产内酯能力较高的菌株,通过感官评价和气相色谱-质谱(gas chromatography-mass spectrometry, GC-MS)技术解析高产脂肪酶乳酸菌及其在不同成熟时间与温度条件下对切达干酪内酯类化合物的贡献,以期为开发改善切达干酪内酯类风味的附属发酵剂以及促进我国干酪产业发展提供理论参考。
二、材料与方法
2.1 材料与试剂
巴氏杀菌乳购自光明乳业股份有限公司;由乳酸乳球菌乳亚种(Lactococcus lactis subsp. lactis)和乳酸乳球菌乳脂亚种(L. lactis subsp. cremoris)组成的Choozit MA 14 LYO发酵剂购自美国国际香精香料公司。乳饼为云南大理市售;臭豆腐卤水为上海浦东新区市售;奶疙瘩为新疆乌鲁木齐市售;奶豆腐为内蒙古阿拉善盟市售。
MARZYME MT2200凝乳酶(米黑根毛霉Rhizomucor miehei酸性蛋白酶,酶活力≥2200 IMCU/g)购自美国国际香精香料公司;MRS肉汤培养基购自北京陆桥技术股份有限公司;正构烷烃(C₇~C₃₀)(色谱纯)购自美国Sigma-Aldrich公司;2-辛醇(分析纯)购自德国Dr. Ehrenstorfer公司;吐温-80、司盘-80(均为分析纯)购自上海阿拉丁生化科技股份有限公司;DNA抽提试剂盒、脂肪酶活性检测试剂盒购自生工生物工程(上海)股份有限公司。
2.2 仪器与设备
Veriti96聚合酶链式反应(polymerase chain reaction, PCR)仪购自美国Applied Biosystems公司;Bio 5000Plus电泳凝胶成像分析系统购自上海中晶科技有限公司;TGL-16M紫外-可见分光光度计购自上海尤尼柯仪器有限公司;DC1741型干酪小型制备机购自上海承欢轻工机械有限公司;DVB/CAR/PDMS固相微萃取纤维头购自美国Supelco公司;7890B-5977B型GC-MS仪购自美国Agilent公司。
2.3 方法
2.3.1 培养基的配制
- 三丁酸甘油酯培养基的配制:100 mL体积分数为10%的三丁酸甘油酯、蛋白胨10 g、牛肉粉3 g、氯化钠5 g、琼脂15 g、900 mL Tris-HCl缓冲液(pH 8.0),121℃灭菌20 min。
- 产酶培养基的配制:20 g橄榄油乳化液(称取1.4 g吐温-80,加入13.2 g去离子水中,边加边搅拌均匀,再于80℃水浴溶解至溶液透明后取出,加入4 g橄榄油和1.4 g司盘-80,用均质机处理20 min至混合液呈乳白色的均匀乳状液,需现用现配)、蛋白胨10 g、牛肉粉10 g、酵母粉5 g、硫酸镁0.1 g、醋酸钠5 g、柠檬酸铵2 g、磷酸二氢钾2 g、硫酸锰0.05 g、蒸馏水1000 mL,pH 7.0,121℃灭菌20 min。
2.3.2 高产脂肪酶乳酸菌的分离筛选与鉴定
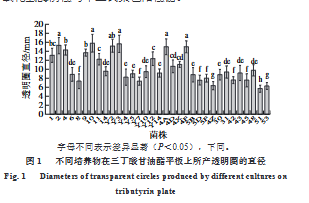
- 产脂肪酶乳酸菌的初筛:在无菌条件下,从4种传统发酵食品(乳饼、臭豆腐卤水、奶疙瘩、奶豆腐)中分离出外观形态不同的单菌落,划线纯化后用30%甘油保存于-80℃。参考李晓楠等[22]的方法并稍作修改,在三丁酸甘油酯平板上打直径为0.8 cm的小孔,精确量取100 μL菌液接种于小孔,置于37℃的培养箱中培养48 h,观察平板上是否出现透明圈,通过测量不同菌株所形成透明圈的直径,初步评估菌株产脂肪酶能力。
- 产脂肪酶乳酸菌的生物学鉴定:将产脂肪酶菌株的单菌落接种于MRS琼脂培养基,37℃培养24 h,观察并记录菌落的大小、形态等特征。挑取单菌落进行过氧化酶实验以及革兰氏染色并镜检,记录菌体形态。对产脂肪酶菌株进行16S rDNA序列鉴定,采用基因组DNA抽提试剂盒提取菌体DNA,PCR扩增引物为27F(5’-GAGAGTTTGATCCTGGCTCAG-3’)和1492R(5’-CGGCTACCTTGTTACGACTT-3’)。PCR扩增条件:95℃预变性10 min;94℃变性30 s、56℃退火30 s、72℃延伸90 s、72℃后延伸5 min,30个循环,最后4℃保存。PCR扩增产物使用1%的琼脂糖凝胶电泳检验,送往生工生物工程(上海)股份有限公司测序。所得16S rDNA的测序结果利用NCBI数据库中的Blast比对分析,将与待鉴定的基因序列同源性大于99%(相似度>99%认定为同一个种[23])的16S rDNA序列使用MEGA11软件构建系统发育树。
- 高产脂肪酶乳酸菌的复筛:测定初筛得到产脂肪酶乳酸菌的脂肪酶活力,筛选高产脂肪酶乳酸菌。将产酶乳酸菌接种于MRS培养基中,置于37℃培养至波长600 nm处的OD值=1.0。随后以2%的接种量分别接种到10 mL产酶培养基中,37℃培养24 h,得到对应的发酵液。取1 mL发酵培养液在7000 r/min、4℃条件下离心10 min,收集上清粗酶液。按照脂肪酶活性检测试剂盒说明书测定,按式(1)计算脂肪酶活力:
[脂肪酶活力 /(U / mL)=frac{A}{t} × F quad (1)]
式中:A为待测样品在710 nm波长处的吸光度;t为催化反应的时间/min;F为发酵液的稀释倍数。
2.3.3 高产脂肪酶乳酸菌的生长特性评价
- 高产脂肪酶乳酸菌耐酸性评价:活化后的高产脂肪酶乳酸菌置于37℃培养至波长600 nm处的OD值=1.0,以2%的接种量分别接种于pH 5.0和pH 6.0的MRS肉汤培养基中,每隔3 h测定不同pH值条件下的OD₆₀₀ₙₘ。
- 高产脂肪酶乳酸菌耐盐性评价:活化后的高产脂肪酶乳酸菌置于37℃培养至波长600 nm处的OD值=1.0,以2%的接种量分别转接于含不同质量分数(2%、3%和5%)NaCl的MRS肉汤培养基中(2%、3%为制备切达干酪通常的NaCl用量[24],考虑到盐分布的均匀性及极端情况下NaCl的质量分数会达到5%[25-26],故将该条件纳入)。于37℃恒温培养箱内培养24 h,每隔3 h测定菌株在不同NaCl含量MRS肉汤培养基中的OD₆₀₀ₙₘ以评价乳酸菌的耐盐性。
- 乳酸菌的自溶度测定:以磷酸盐缓冲液为对照,测定菌体悬液和对照在37℃条件下培养24 h的OD₆₀₀ₙₘ。按式(2)计算菌体的自溶度:
[自溶度 / \%=frac{OD_{0}-OD_{24}}{OD_{0}} × 100 quad (2)]
式中:OD₀为初始的OD值;OD₂₄为培养24 h后的OD值。
2.3.4 附属发酵剂的制备
取生长特性较好的高产脂肪酶乳酸菌接种于10 mL MRS肉汤培养基中,37℃恒温培养至波长600 nm处的OD值=1.0。按2%的接种量转接于50 mL产酶培养基中,37℃恒温培养24 h后取上述发酵液到离心管中,在4℃、10000 r/min条件下离心10 min,沉淀物用质量分数0.85%生理盐水洗涤2次,最后将菌泥重悬于0.85%生理盐水中得到附属发酵剂菌液。
2.3.5 干酪浆快速成熟模型的制作
参考郭婷等[20]的方法。巴氏杀菌乳水浴加热至30℃后测定pH值(pH₁),然后向巴氏杀菌乳中加入1%(质量分数)的商业发酵剂(乳酸乳球菌乳亚种和乳酸乳球菌乳脂亚种),并在30℃条件下发酵,在发酵过程中测定pH值(pH₂),当pH₁与pH₂间差值大于0.4时,向其中加入1.5%(质量分数)的凝乳酶,搅拌混匀后静置40 min。等待巴氏杀菌乳凝固后,将其切割成豆腐乳样形状,并在切割过程中继续水浴加热,以0.2℃/min的速率升温至38℃,待析出乳清的pH值降至6.15时,排出乳清并保持温度为38℃,当凝乳凝结成块后将其切割成30 cm×30 cm×30 cm的块状。每隔15~20 min翻转1次,重复3次,待排出的乳清pH值降至5.45时完成凝块的堆积,得到干酪凝块。随后取100 g干酪凝块装入无菌真空袋中,对照组加入50 mL质量分数为5%的灭菌NaCl溶液,实验组加入47 mL质量分数为5%的灭菌NaCl溶液和3 mL附属发酵剂菌液(乳酸菌浓度为10×10⁹ CFU/mL,添加量0.8%)搅打成浆状,真空密封完成干酪浆快速成熟模型的制作。在30℃恒温环境中成熟1、3、6、9、12 d。根据干酪浆中内酯化合物的含量,筛选出产内酯性能较好的高产脂肪酶乳酸菌。
2.3.6 切达干酪的制备
参考本课题组前期建立的方法[3]。巴氏杀菌乳打入干酪罐中并加热至31℃,记录此时的pH值,分别加入质量分数1%发酵剂(乳酸乳球菌乳亚种和乳酸乳球菌乳脂亚种)和附属发酵剂(乳酸菌浓度为1×10¹⁰ CFU/mL,添加量0.8%),并设置未添加附属发酵剂的对照组,随后在31℃条件下发酵30 min,待杀菌乳pH值与发酵前相比下降0.4时加入质量分数1.5%凝乳酶,搅拌后凝乳40 min。随后将凝块切割成边长为1 cm的方块,切割过程中以0.1℃/min的速率热烫至39℃,随后将干酪罐盖上盖子使温度维持在39℃,待析出乳清的pH值为6.15时排出乳清,将凝乳切割成30 cm×30 cm×30 cm的方块。干酪块堆积在一起,每隔15 min翻转1次,重复3次。待析出乳清的pH值为5.45后将凝乳块切成1 cm×1 cm×1 cm的方块。随后将质量分数2.5%的NaCl溶液均匀撒到切达干酪上并进行搅拌,入模型压榨成型并真空包装,分别放于4、10、14℃(10~14℃有助于加快干酪的成熟[3,30])恒温条件成熟30、60、90、120、150 d(切达干酪的成熟期为2~36个月[30-31])。
2.3.7 干酪的感官评价
由20名经感官品评培训的评价员组成评价小组(女性10名、男性10名,平均年龄24岁)。所有的感官测试按照国际标准ISO 8589:2007在感官实验室中执行,室内温度控制在20℃。待测试的干酪样品用数字随机编码,每个干酪样品切成大小均匀的颗粒并称取5 g,分别保存在一个有盖的、无气味的玻璃容器中(总容量为50 mL)。然后,按随机顺序排列,依次呈送给评价员进行感官评价测试。感官评价员按照评价表上的属性和定义对每个干酪样品进行评分,所采用的描述词与参考标准分别为奶香味(0.002%的丁二酮溶液)、果香味(0.002%的己酸乙酯溶液)、硫味(捣碎的鸡蛋壳)、酸腐味(0.1%的丁酸溶液)、坚果味(生坚果)、干酪味(新鲜黄油)。在干酪香气感官评价中,每个香气属性以0~10进行强度等级划分(0为无强度或香气无察觉,5为中等强度,10为非常强)。每个干酪样品重复检验3次。
2.3.8 干酪的GC-MS分析
采用顶空固相微萃取法对干酪中内酯类化合物进行提取。分别精确称量5.0 g干酪样品,碾碎后放入萃取瓶中,加入100 μL 220 mg/L的2-辛醇溶液,在60℃的恒温水浴锅中平衡5 min,通过DVB/CAR/PDMS固相微萃取头在60℃条件下萃取45 min。采用HP-INNOWAX色谱柱(60 m×0.25 mm,0.25 μm);进样口温度250℃。程序升温:40℃保持3 min,以5℃/min的速率升至140℃,保持2 min,3℃/min升至240℃,保持15 min。载气为氦气(纯度为99.99%);流量为2 mL/min;进样方式为不分流进样。
质谱条件:电子轰击能量:70 eV;离子源温度:230℃;四极杆温度:150℃;发射电流:35 µA;扫描速率:1.9 scans/s;质量扫描范围m/z 30~450。所有挥发性化合物的定性采用NIST17质谱库进行检索,选择匹配度大于80%的物质确定化合物。再利用正构烷烃C₇~C₃₀计算保留指数(retention index, RI),并与文献报道值进行比对,两者结合对干酪内酯类化合物进行定性分析。采用内标法进行定量分析,分别按式(3)、(4)计算各组分含量和RI:
[内酯类化合物含量 /(mg / kg)=frac{
ho_{0} × V_{0}}{m} × frac{S_{a}}{S_{0}} quad (3)]
式中:ρ₀为内标物的质量浓度/(mg/L);V₀为内标物的体积/L;m为样品质量/kg;Sₐ为待测化合物的峰面积;S₀为内标物的峰面积。
[RI=100 × n+100 × frac{RT-RT_{n}}{RT_{n+1}-RT_{n}} quad (4)]
式中:RT为化合物的保留时间/min;RTₙ和RTₙ₊₁分别为碳数n和n+1正构烷烃的保留时间/min(RTₙ
2.4 数据处理
每个实验重复3次,采用Excel 2019软件整理实验数据,以±s表示。采用SPSS 26.0软件对数据进行显著性分析,利用Origin 2019b软件绘图。
三、结果与分析
3.1 高产脂肪酶乳酸菌的筛选
三丁酸甘油酯平板上培养物周围出现透明圈表明该培养物可产生降解三丁酸甘油酯(C₄)中短链底物的脂肪酶,因此该方法常用于脂肪酶生产菌的筛选[32]。本研究采用三丁酸甘油酯透明圈法从乳饼、臭豆腐卤水、奶疙瘩、奶豆腐中筛选出33株具有透明圈形成能力的培养物。将初筛培养物经划线纯化后在MRS琼脂平板上的菌落形态呈白色或乳白色,菌落较小且呈圆形,边缘光滑。经过过氧化氢酶实验与革兰氏染色得到10株过氧化氢酶阴性与革兰氏染色阳性菌。
将菌株的16S rDNA序列上传至NCBI数据库进行Blast比对,选取相似性大于99%的菌株序列,使用MEGA11软件将其与分离菌株进行多序列比对,构建系统发育树。结果显示,菌株Y3、Y2为格氏乳球菌(L. garvieae),4X、42、6为戊糖片球菌(Pediococcus pentosaceus),4D、2为乳酸片球菌(P. acidilactici),31、43、30为植物乳植杆菌(Lactiplantibacillus plantarum)。
随后利用铜皂法对初筛具有产脂肪酶能力的乳酸菌所产脂肪酶的活性进行测定,结果显示,从云南、新疆、内蒙古等主要牧民生活地区发酵乳制品所分离的戊糖片球菌6、戊糖片球菌4X、乳酸片球菌4D、乳酸片球菌2所产脂肪酶具有相对较高的活力,其中乳酸片球菌4D酶活力最高,达3.32±0.54 U/mL,格氏乳球菌Y3酶活力为1.46±0.27 U/mL。
3.2 高产脂肪酶菌株的生长特性分析
3.2.1 耐酸性与耐盐性分析
研究菌株生长环境中NaCl含量与pH值对其生长的影响情况对于切达干酪的制备具有重要意义。在切达干酪成熟过程(60~180 d)中的pH值范围通常在5.0~6.0之间[33]。在pH 5.0、6.0的MRS培养基中,6株产脂肪酶乳酸菌均可正常生长。在2%、3% NaCl溶液环境中6株产酶乳酸菌均能够正常生长;当NaCl质量分数达到5%时,格氏乳球菌Y2和戊糖片球菌6的生长受到抑制。这表明格氏乳球菌Y3、乳酸片球菌2、戊糖片球菌4X和乳酸片球菌4D能够较好适应常规以及非常规切达干酪的环境,也可用于干酪浆快速成熟模型的制作,从而进一步评估菌株在干酪中的应用潜力。
3.2.2 菌株的自溶度分析
菌株的自溶度是影响干酪质量和口味的重要因素,它可以帮助细菌产生胞内酶,从而改善干酪的风味[34-35]。戊糖片球菌4X、格氏乳球菌Y3、乳酸片球菌4D、乳酸片球菌2的自溶度均大于15%。研究表明,菌株的自溶度大于15%能够快速释放胞内酶进行代谢反应,从而促进风味化合物的形成[36-37]。综合不同产脂肪酶乳酸菌的耐酸耐盐特性,选择戊糖片球菌4X、格氏乳球菌Y3、乳酸片球菌4D、乳酸片球菌2用于建立干酪浆快速成熟模型,分析其产内酯能力。
3.3 基于干酪浆快速成熟模型的高产脂肪酶乳酸菌产内酯化合物性能分析
在干酪浆的12 d成熟期内,由不同产脂肪酶乳酸菌发酵的干酪浆中内酯总含量在成熟的第3天到第9天迅速增加,在成熟后期又缓慢增加;乳酸片球菌4D发酵成熟的干酪浆中总内酯含量(181.88 μg/kg)高于其他不同产酶乳酸菌发酵干酪浆,且是对照组样品(71.25 μg/kg)的2.6倍,格氏乳球菌Y3发酵的干酪浆中总内酯含量(157.75 μg/kg)是对照组的2.2倍。在5组干酪浆样品中均鉴定出δ-癸内酯和δ-十二内酯;在乳酸片球菌4D和格氏乳球菌Y3发酵的干酪浆中还鉴定出δ-辛内酯,第12天的含量分别为25.78 μg/kg和39.71 μg/kg;仅在格氏乳球菌Y3发酵的干酪浆中鉴定出γ-丁内酯(84.1 μg/kg)。本研究发现乳酸片球菌4D和格氏乳球菌Y3能够丰富干酪浆快速成熟模型中的内酯化合物种类与含量,具有促进内酯风味形成的潜力,因此选择这两株乳酸菌进一步探究产脂肪酶菌株对切达干酪产品风味的影响作用。
3.4 不同高产脂肪酶乳酸菌所制备干酪的感官评价
干酪的成熟温度与成熟时间会改变菌株的代谢活动,从而影响干酪的风味品质[38-39]。通过对分别添加了2株产脂肪酶乳酸菌和未添加产脂肪酶乳酸菌的干酪香气进行感官评价,探究其在4、10℃和14℃条件下成熟30、60、90、120、150 d的风味变化情况。感官评价结果表明乳酸片球菌4D和格氏乳球菌Y3发酵干酪样品的风味强度随时间的延长、温度的增加而逐渐增强。在10、14℃成熟90 d时,添加高产脂肪酶乳酸菌4D和Y3干酪样品的奶香味和果香味均高于对照组,且在14℃条件下添加乳酸片球菌4D干酪的奶香味和果香味强度值达到最高,分别为7.9和7.4。而含有格氏乳球菌Y3的干酪在14℃成熟120 d时,具有最高的奶香味香气强度,为7.9。在成熟150 d时,不同成熟温度下的干酪风味强度均降低。
3.5 不同成熟时间和温度下高产脂肪酶乳酸菌所制备干酪的内酯类化合物分析
添加2种高产脂肪酶乳酸菌以及未添加高产脂肪酶乳酸菌的切达干酪在150 d成熟周期中内酯化合物种类以及含量变化显示,通过NIST 17.0谱库与RI共检出4种内酯类化合物,包括δ-癸内酯、δ-十二内酯、δ-辛内酯和γ-丁内酯。δ-癸内酯是切达干酪中最为常见的内酯化合物,赋予干酪椰子的风味属性[40]。对照组干酪的δ-癸内酯含量最高达106.05 μg/kg,添加乳酸片球菌4D和格氏乳球菌Y3的干酪最高分别可达245.33、353.24 μg/kg。δ-十二内酯通常赋予干酪奶油以及奶香的风味,已有研究表明其是干酪中的关键风味化合物[41]。δ-十二内酯在对照组成熟过程中含量最高达44.58 μg/kg,添加乳酸片球菌4D与格氏乳球菌Y3的干酪中δ-十二内酯的最高含量分别为108.85、77.17 μg/kg。δ-辛内酯也是干酪中的主要内酯,其对干酪的风味有重要贡献,能够赋予干酪椰子的风味属性[42-43]。添加乳酸片球菌4D与格氏乳球菌Y3的干酪在成熟前60 d即可检测到δ-辛内酯,而对照组在成熟90 d后才能够检测出。此外,仅在添加格氏乳球菌Y3的干酪样品中检测到γ-丁内酯,其可贡献椰子味和奶油味的风味属性[44]。综上,产脂肪酶乳酸菌能够提高干酪中内酯化合物的含量与种类,且在所筛选的两株高产脂肪酶乳酸菌中,格氏乳球菌Y3所发酵的干酪含有较高含量的内酯化合物与种类,因此其具有的产内酯性能更好。
影响切达干酪中内酯化合物形成的因素包括温度、水分、附属发酵剂以及原料乳的预处理方式[45]。本研究探究了不同成熟时间和温度对干酪样品中内酯化合物形成的影响。结果表明,在4~14℃成熟120 d时,切达干酪中的δ-癸内酯含量最高,δ-辛内酯与δ-十二内酯均在成熟90 d时检测到最高值,且内酯含量随成熟温度的提高而提高。γ-丁内酯在4~10℃成熟150 d时含量达最高(24.52~38.45 μg/kg),而在14℃成熟90 d时可达到最高(54.25 μg/kg)。可见,较高的成熟温度能够促进内酯化合物的形成。此外,在14℃成熟120 d后干酪中内酯化合物含量逐渐降低或积累变慢。因此,较高的成熟温度虽然对干酪内酯风味具有积极影响,但成熟时间不宜过长。综上,选择产脂肪酶乳酸菌Y3发酵干酪,且在14℃成熟120 d适于干酪内酯化合物的形成。
四、讨论
切达干酪中的内酯类化合物是牛乳中甘油三酯在脂肪酶的作用下形成脂肪酸,进一步在β-氧化酶、环化酶的作用下形成,对干酪整体风味具有重要贡献[46]。研究发现,产脂肪酶乳酸乳球菌乳亚种(双乙酰型)(L. lactis subsp. lactis biovar diacetylactis)能够提高干酪中脂肪酸的含量[47]。因此,将产脂肪酶乳酸菌作为附属发酵剂与商业发酵剂混合可作为调控干酪内酯类化合物生物合成的策略。干酪浆快速成熟模型因具有较高的含水量与成熟温度,可用于快速评估附属发酵剂乳酸菌的风味形成潜力[17]。此外,探究成熟温度及时间对切达干酪风味形成的影响已成为一个热点[3,28,48],但目前对于产脂肪酶乳酸菌与切达干酪内酯化合物之间的相关性鲜有报道。本研究基于三丁酸甘油酯平板、铜皂法和干酪浆快速成熟模型筛选适用于制备切达干酪、具有较好产内酯能力的高产脂肪酶乳酸菌并制备附属发酵剂,结合感官评价与GC-MS探究含高产脂肪酶乳酸菌的附属发酵剂以及不同成熟温度与时间对切达干酪内酯化合物形成的影响。
脂肪酶能够催化甘油三酯的水解从而形成切达干酪的风味前体——游离脂肪酸,通常外源添加的脂肪酶适用于原料乳,因此利用附属发酵剂在干酪发酵过程中产生脂肪酶,可避免因原料乳热处理引起的酶失活[49]。乳酸菌是脂肪酶的来源之一,如Esteban-Torres等[49]发现植物乳植杆菌所产的脂肪酶能够降解三丁酸甘油酯。本研究从发酵乳制品所分离的戊糖片球菌6、戊糖片球菌4X、乳酸片球菌4D、乳酸片球菌2具有相对较高的脂肪酶活力。李晓楠等[22]结合三丁酸甘油酯平板透明圈法以及铜皂法从新疆地区发酵乳制品中筛选出高产脂肪酶的植物乳植杆菌、瑞士乳酸杆菌(L. helveticus)。此外,从臭豆腐卤水分离出的格氏乳球菌Y2、格氏乳球菌Y3所产的脂肪酶活性也相对较高。有研究表明臭豆腐卤水中含有较多微生物所产生的脂肪酶[50]。目前研究的产脂肪酶菌株主要是从土壤、污水等环境中筛选分离[51-52],本研究结果为食源产脂肪酶乳酸菌的筛选提供了新的参考。
干酪浆快速成熟模型与切达干酪在NaCl添加量方面存在差异,前者的NaCl添加量通常在5%,切达干酪中的NaCl质量分数在2%~4%[24,27]。结合不同NaCl浓度下的生长曲线,发现格氏乳球菌Y3、乳酸片球菌2、戊糖片球菌4X和乳酸片球菌4D在5%的NaCl环境下能够生长,因此可应用于干酪浆快速成熟模型。干酪浆快速成熟模型能够缩短干酪的成熟周期[17-19]。Kristoffersen等[27]发现干酪浆在30℃成熟3 d后,可形成切达干酪的独特风味。将格氏乳球菌Y3、乳酸片球菌2、戊糖片球菌4X和乳酸片球菌4D应用于干酪浆快速成熟模型,发现在干酪浆12 d的成熟期中,内酯化合物含量从第3天开始迅速增加。这一加速过程是由于干酪浆具有较高的含水量和成熟温度,从而加快体系内微生物的生化代谢[27]。但有研究表明,这种高温、高水分的环境会影响体系中的化学反应和酶催化反应以及增加微生物腐败等风险,因此干酪浆快速模型更适合筛选特定的微生物[53]。另一方面,乳酸菌的自溶与风味密切相关,如Drake等[54]通过促进发酵剂菌株的自溶改善干酪品质。乳酸片球菌4D和格氏乳球菌Y3的自溶度与其他菌株相比相对较高,这可能与其较好的产内酯性能相关。
附属发酵剂的添加对切达干酪的感官评分与风味形成具有重要的影响[38]。将干酪浆快速成熟模型筛选出产内酯性能较好的乳酸片球菌4D和格氏乳球菌Y3制成附属发酵剂,并应用于切达干酪的制备,发现添加高产脂肪酶乳酸菌4D和Y3干酪样品具有更浓郁的奶香味和果香味。添加乳酸片球菌4D和格氏乳球菌Y3提高了切达干酪中δ-癸内酯、δ-十二内酯的含量,且缩短了δ-辛内酯的形成时间,只有格氏乳球菌Y3在成熟过程中形成γ-丁内酯。研究表明,乳球菌能够促进脂肪分解并有效改善干酪风味[55]。李宇辉等[56]发现产脂肪酶乳酸菌附属发酵剂能够改善干酪的感官品质,促进具有奶油味、果香味的风味化合物形成。因此产脂肪酶乳酸菌能够改善切达干酪的风味感官,促进内酯类化合物的形成。
成熟温度对切达干酪的风味也有影响[43]。切达干酪中的内酯化合物含量随成熟温度的提高呈上升趋势,其中14℃相比其他2种温度能够加快内酯化合物的形成,如γ-丁内酯在4~10℃成熟150 d时含量达到最高,而在14℃成熟90 d即可达到最高值。这可能是由于干酪中微生物菌群的代谢酶活性被激活,从而促进微生物代谢产香[57-58]。Walsh等[30]发现10℃成熟8个月与7℃成熟12个月的干酪感官品质相似,较高的成熟温度能够促进干酪风味的形成,这与本研究结果一致。另一方面,成熟时间对内酯的积累具有重要作用,Alewijn等[59]发现γ-内酯类化合物需要更长的成熟时间才能达到最高浓度,这与本研究中γ-丁内酯的积累趋势一致。王姣等[48]对不同成熟周期切达干酪的挥发性化合物分析发现,切达干酪中内酯化合物的含量随成熟时间的延长缓慢增加。Chen Chen等[8]对英国威克农场切达干酪的内酯化合物进行分析,发现δ-辛内酯、δ-癸内酯以及δ-十二内酯在早期至成熟阶段迅速积累,随后缓慢变化。在添加产脂肪酶乳酸菌附属发酵剂的切达干酪中,这3种内酯化合物在成熟90~120 d达到最高值,随后逐渐下降。但对于内酯化合物与切达干酪的成熟标志之间相关性,还需通过风味指纹图谱技术结合多元统计方法进一步分析判断。
五、结论
本研究利用三丁酸甘油酯透明圈法、铜皂法并结合干酪浆快速成熟模型筛选得到耐酸耐盐性与产内酯性能较好的乳酸片球菌4D和格氏乳球菌Y3。将乳酸片球菌4D和格氏乳球菌Y3作为附属发酵剂用于切达干酪的制备,研究发现添加这两株菌的切达干酪中内酯化合物种类与含量均提高,且改善了切达干酪的感官属性。格氏乳球菌Y3能够促进切达干酪γ-丁内酯的形成,且在14℃成熟120 d的效果较好,在切达干酪的附属发酵剂开发方面具有良好的应用潜力。后续研究可进一步测定干酪成熟过程中脂肪酸含量的变化,以探究乳酸菌所产脂肪酶在内酯化合物形成过程中的作用以及内酯化合物的形成与切达干酪成熟的相关性。
参考文献
[1] KENDIRCI P, SALUM P, BAS D, et al. Production of enzyme-modified cheese (EMC) with ripened white cheese flavour: Ⅱ-effects of lipases[J]. Food and Bioproducts Processing, 2020, 122: 230-244. DOI:10.1016/j.fbp.2020.05.010.
[2] CHEN C, TIAN T H, YU H Y, et al. Characterisation of the key volatile compounds of commercial Gouda cheeses and their contribution to aromas according to Chinese consumers’ preferences[J]. Food Chemistry: X, 2022, 15: 100416. DOI:10.1016/j.fochx.2022.100416.
[3] 陈臣, 田同辉, 周文雅, 等. 高产3-甲基丁醛乳酸乳球菌对切达干酪风味的影响[J]. 中国食品学报, 2023, 23(11): 276-288. DOI:10.16429/j.1009-7848.2023.11.027.
[4] XIA A N, YANG Y, GUO M, et al. Unveiling of the key pathway in flavor formation in fermented milk of Lactococcus lactis subsp. lactis via genomics and metabolomics[J]. Food Bioscience, 2023, 56: 103159. DOI:10.1016/j.fbio.2023.103159.
[5] JIANG N, WU R N, WU C, et al. Multi-omics approaches to elucidate the role of interactions between microbial communities in cheese flavor and quality[J]. Food Reviews International, 2023, 39(8): 5446-5458. DOI:10.1080/87559129.2022.2070199.
[6] GOBBETTI M, DE ANGELIS M, DI CAGNO R, et al. Pros and cons for using non-starter lactic acid bacteria (NSLAB) as secondary/adjunct starters for cheese ripening[J]. Trends in Food Science & Technology, 2015, 45(2): 167-178. DOI:10.1016/j.tifs.2015.07.016.
[7] 康优, 吕杨, 高磊, 等. 乳酸片球菌与马克斯克鲁维酵母混合附属发酵剂对切达干酪品质的影响[J/OL]. 食品科学, 2024 (2024-09-19). https://kns.cnki.net/kcms/detail/11.2206.TS.20240918.0959.039.html.
[8] CHEN C, LIU Z, YU H Y, et al. Flavoromic determination of lactones in cheddar cheese by GC-MS-olfactometry, aroma extract dilution analysis, aroma recombination and omission analysis[J]. Food Chemistry, 2022, 368: 130736. DOI:10.1016/j.foodchem.2021.130736.
[9] SUTAY KOCABAŞ D, LYNE J, USTUNOL Z. Hydrolytic enzymes in the dairy industry: applications, market and future perspectives[J]. Trends in Food Science & Technology, 2022, 119: 467-475. DOI:10.1016/j.tifs.2021.12.013.
[10] BORA L, GOHAIN D, DAS R. Recent advances in production and biotechnological applications of thermostable and alkaline bacterial lipases[J]. Journal of Chemical Technology & Biotechnology, 2013, 88(11): 1959-1970. DOI:10.1002/jctb.4170.
[11] HUANG Y Y, YU J J, ZHOU Q Y, et al. Preparation of yogurt-flavored bases by mixed lactic acid bacteria with the addition of lipase[J]. LWT-Food Science and Technology, 2020, 131: 109577. DOI:10.1016/j.lwt.2020.109577.
[12] CARON T, LE PIVER M, PÉRON A C, et al. Strong effect of Penicillium roqueforti populations on volatile and metabolic compounds responsible for aromas, flavor and texture in blue cheeses[J]. International Journal of Food Microbiology, 2021, 354: 109174. DOI:10.1016/j.ijfoodmicro.2021.109174.
[13] RANI S, JAGTAP S. Acceleration of Swiss cheese ripening by microbial lipase without affecting its quality characteristics[J]. Journal of Food Science and Technology, 2019, 56(1): 497-506. DOI:10.1007/s13197-018-3482-6.
[14] 杨万龄, 沈兴旺, 崔凤怡, 等. 蓝纹牦牛干酪成熟特性及其风味分析[J]. 食品科学技术学报, 2023, 41(6): 52-64. DOI:10.12301/spxb202300008.
[15] XIANG Q, XIA Y X, CHEN L, et al. Flavor precursors and flavor compounds in Cheddar-flavored enzyme-modified cheese due to pre-enzymolysis combined with lactic acid bacteria fermentation[J]. Food Bioscience, 2023, 53: 102698. DOI:10.1016/j.fbio.2023.102698.
[16] RANDAZZO C L, LIOTTA L, ANGELIS M, et al. Adjunct culture of non-starter lactic acid bacteria for the production of provola Dei nebrodi PDO cheese: in vitro screening and pilot-scale cheesemaking[J]. Microorganisms, 2021, 9(1): 179. DOI:10.3390/microorganisms9010179.
[17] CASTADA H Z, HARPER W J, BARRINGER S A. Volatile organic compounds of a Swiss cheese slurry system with and without added reduced glutathione, compared with commercial Swiss cheese[J]. International Dairy Journal, 2015, 49: 72-77. DOI:10.1016/j.idairyj.2015.04.010.
[18] DARNAY L, MIKLÓS G, LŐRINCZ A, et al. Possible inhibitory effect of microbial transglutaminase on the formation of biogenic amines during Trappist cheese ripening[J]. Food Additives & Contaminants. Part A, Chemistry, Analysis, Control, Exposure & Risk Assessment, 2022, 39(3): 580-587. DOI:10.1080/19440049.2021.2005831.
[19] BORAN O S, SULEJMANI E, HAYALOGLU A A. Acceleration of proteolysis, flavour development and enhanced bioactivity in a model cheese using Kuflu cheese slurry: an optimisation study[J]. Food Chemistry, 2023, 412: 135495. DOI:10.1016/j.foodchem.2023.135495.
[20] 郭婷, 余志坚, 陈超, 等. 基于快速成熟模型的藏灵菇发酵切达干酪挥发性风味物质分析[J]. 食品科学, 2018, 39(8): 90-96. DOI:10.7506/spkx1002-6630-201808015.
[21] 李春燕, 熊智强, 王光强, 等. 乳酸菌附属发酵剂的筛选及其对干酪浆蛋白水解的影响[J]. 食品与发酵工业, 2022, 48(17): 129-135. DOI:10.13995/j.cnki.11-1802/ts.030574.
[22] 李晓楠, 李宇辉, 卢士玲, 等. 新疆传统奶酪中产脂肪酶乳酸菌的优选及酶学特性研究[J]. 食品与发酵工业, 2021, 47(10): 50-57. DOI:10.13995/j.cnki.11-1802/ts.025978.
[23] 李斯淼, 陈芳敏, 方丽, 等. 一株家蚕肠道分离乳酸菌的鉴定及生物学特性[J/OL]. 微生物学通报. https://doi.org/10.13344/j.microbiol.china.240512.
[24] 吴怡霏, 刘宗尚, 刘维娜, 等. NaCl对原制干酪品质影响及减钠方法研究进展[J]. 中国食品添加剂, 2022, 33(5): 186-193. DOI:10.19804/j.issn1006-2513.2022.05.025.
[25] ONG L, D’INCECCO P, PELLEGRINO L, et al. The effect of salt on the structure of individual fat globules and the microstructure of dry salted Cheddar cheese[J]. Food Biophysics, 2020, 15(1): 85-96. DOI:10.1007/s11483-019-09606-x.
[26] MISTRY V V, KASPERSON K M. Influence of salt on the quality of reduced fat Cheddar cheese 1[J]. Journal of Dairy Science, 1998, 81(5): 1214-1221. DOI:10.3168/jds.S0022-0302(98)75681-4.
[27] KRISTOFFERSEN T, MIKOLAJCIK E M, GOULD I A. Cheddar cheese flavor. IV. directed and accelerated ripening process[J]. Journal of Dairy Science, 1967, 50(3): 292-297. DOI:10.3168/jds.s00220302(67)87412-5.
[28] 莫蓓红, 赵建, 苗君莅, 等. 一种植物乳杆菌对干酪模型成熟中微生物及蛋白质水解的影响[J]. 食品工业科技, 2012, 33(7): 100-104. DOI:10.13386/j.issn1002-0306.2012.07.062.
[29] MUEHLENKAMP-ULATE M R, WARTHESEN J J. Evaluation of several nonstarter lactobacilli for their influence on Cheddar cheese slurry proteolysis 1[J]. Journal of Dairy Science, 1999, 82(7): 1370-1378. DOI:10.3168/jds.S0022-0302(99)75362-2.
[30] WALSH E A, DIAKO C, SMITH D M, et al. Influence of storage time and elevated ripening temperature on the chemical and sensory properties of white Cheddar cheese[J]. Journal of Food Science, 2020, 85(2): 268-278. DOI:10.1111/1750-3841.14998.
[31] 程晶晶, 陈会民, 罗洁, 等. 不同成熟期切达干酪的组分及特征风味物质的解析[J]. 食品与发酵工业, 2020, 46(15): 231-237. DOI:10.13995/j.cnki.11-1802/ts.023149.
[32] GARCÍA-CANO I, ROCHA-MENDOZA D, KOSMERL E, et al. Purification and characterization of a phospholipid-hydrolyzing phosphoesterase produced by Pediococcus acidilactici isolated from Gouda cheese[J]. Journal of Dairy Science, 2020, 103(5): 3912-3923. DOI:10.3168/jds.2019-17965.
[33] XIA Y J, YUAN R, WENG S N, et al. Proteolysis, lipolysis, texture and sensory properties of cheese ripened by Monascus fumeus[J]. Food Research International, 2020, 137: 109657. DOI:10.1016/j.foodres.2020.109657.
[34] ZHAO S Q, DORAU R, TØMMERHOLT L, et al. Simple & better-accelerated cheese ripening using a mesophilic starter based on a single strain with superior autolytic properties[J]. International Journal of Food Microbiology, 2023, 407: 110398. DOI:10.1016/j.ijfoodmicro.2023.110398.
[35] KANG O J, VÉZINZ L P, LABERGE S, et al. Some factors influencing the autolysis of Lactobacillus bulgaricus and Lactobacillus casei[J]. Journal of Dairy Science, 1998, 81(3): 639-646. DOI:10.3168/jds.s0022-0302(98)75618-8.
[36] AYAD E H E, NASHAT S, EL-SADEK N, et al. Selection of wild lactic acid bacteria isolated from traditional Egyptian dairy products according to production and technological criteria[J]. Food Microbiology, 2004, 21(6): 715-725. DOI:10.1016/j.fm.2004.02.009.
[37] TONDHOUSH A, SOLTANI M, AZARIKIA F, et al. Fabrication of UF-white cheese: Obtaining a different proteolysis rate, texture, and flavor via using combinations of mesophilic starter culture and Lactobacillus helveticus[J]. Food Science & Nutrition, 2023, 12(1): 328-339. DOI:10.1002/fsn3.3769.
[38] BASSI D, GAZZOLA S, SATTIN E, et al. Lactic acid bacteria adjunct cultures exert a mitigation effect against spoilage microbiota in fresh cheese[J]. Microorganisms, 2020, 8(8): 1199. DOI:10.3390/microorganisms8081199.
[39] SIHUFE G A, ZORRILLA S E, PEROTTI M C, et al. Acceleration of cheese ripening at elevated temperature. an estimation of the optimal ripening time of a traditional Argentinean hard cheese[J]. Food Chemistry, 2010, 119(1): 101-107. DOI:10.1016/j.foodchem.2009.06.001.
[40] DELGADO F J, GONZÁLEZ-CRESPO J, CAVA R, et al. Formation of the aroma of a raw goat milk cheese during maturation analysed by SPME-GC-MS[J]. Food Chemistry, 2011, 129(3): 1156-1163. DOI:10.1016/j.foodchem.2011.05.096.
[41] HIGH R, BREMER P, KEBEDE B, et al. Comparison of four extraction techniques for the evaluation of volatile compounds in spray-dried New Zealand sheep milk[J]. Molecules, 2019, 24(10): 1917. DOI:10.3390/molecules24101917.
[42] PLUTA-KUBICA A, DOMAGAŁA J, GĄSIOR R, et al. Characterisation of the profile of volatiles of Polish Emmental cheese[J]. International Dairy Journal, 2021, 116: 104954. DOI:10.1016/j.idairyj.2020.104954.
[43] QIAN M, REINECCIUS G. Identification of aroma compounds in Parmigiano-Reggiano cheese by gas chromatography/olfactometry[J]. Journal of Dairy Science, 2002, 85(6): 1362-1369. DOI:10.3168/jds.S0022-0302(02)74202-1.
[44] FAN S S, TANG K, XU Y, et al. Characterization of the potent odorants in Tibetan Qingke Jiu by sensory analysis, aroma extract dilution analysis, quantitative analysis and odor activity values[J]. Food Research International, 2020, 137: 109349. DOI:10.1016/j.foodres.2020.109349.
[45] 于海燕, 姚文倩, 陈臣, 等. 内酯类化合物在食品中的风味贡献及形成机制[J]. 现代食品科技, 2022, 38(5): 337-349; 55. DOI:10.13982/j.mfst.1673-9078.2022.5.0954.
[46] 陈臣, 刘政, 于海燕, 等. 奶酪中内酯类物质风味贡献及其生物合成调控进展[J]. 现代食品科技, 2020, 36(11): 305-312. DOI:10.13982/j.mfst.1673-9078.2020.11.0513.
[47] TUNGJAROENCHAI W, WHITE C H, HOLMES W E, et al. Influence of adjunct cultures on volatile free fatty acids in reduced-fat Edam cheeses[J]. Journal of Dairy Science, 2004, 87(10): 3224-3234. DOI:10.3168/jds.S0022-0302(04)73458-X.
[48] 王姣, 许凌云, 张晋华, 等. 不同成熟时间切达奶酪中挥发性香气成分及其电子鼻判别分析[J]. 食品科学, 2020, 41(20): 175-183. DOI:10.7506/spkx1002-6630-20190726-349.
[49] ESTEBAN-TORRES M, MANCHEÑO J M, DE LAS RIVAS B, et al. Production and characterization of a tributyrin esterase from Lactobacillus plantarum suitable for cheese lipolysis[J]. Journal of Dairy Science, 2014, 97(11): 6737-6744. DOI:10.3168/jds.2014-8234.
[50] TIAN H X, ZOU L, LI L, et al. Characterisation of the aroma profile and dynamic changes in the flavour of stinky tofu during storage[J]. Foods, 2023, 12(7): 1410. DOI:10.3390/foods12071410.
[51] 许琳琳, 刘慧乾, 张梦瑶, 等. 低温脂肪酶产生菌的筛选、表达及酶学性质分析[J]. 食品工业科技, 2024, 45(20): 133-140. DOI:10.13386/j.issn1002-0306.2023110093.
[52] 王娜, 赵文倩, 张盈盈, 等. 产脂肪酶多粘类芽胞杆菌的产酶条件和固定化优化及对模拟油污废水的降解研究[J]. 微生物学杂志, 2024, 44(4): 31-39. DOI:10.3969/j.issn.1005-7021.2024.04.004.
[53] FOX P F, WALLACE J M, MORGAN S, et al. Acceleration of cheese ripening[J]. Antonie van Leeuwenhoek, 1996, 70(2/3/4): 271-297. DOI:10.1007/BF00395937.
[54] DRAKE M A, BOYLSTON T D, SPENCE K D, et al. Chemical and sensory effects of a Lactobacillus adjunct in Cheddar cheese[J]. Food Research International, 1996, 29(3/4): 381-387. DOI:10.1016/0963-9969(96)00006-3.
[55] KLEIN N, LORTAL S. Attenuated starters: an efficient means to influence cheese ripening: a review[J]. International Dairy Journal, 1999, 9(11): 751-762. DOI:10.1016/S0958-6946(99)00155-7.
[56] 李宇辉, 王俊钢, 刘成江, 等. 产脂肪酶乳酸菌对新疆传统奶酪脂肪酸及风味的影响[J]. 农业工程学报, 2022, 38(6): 319-329.
[57] UEDA S, SASAKI R, NAKABAYASHI R, et al. Exploring the lipids involved in the formation of characteristic lactones in Japanese black cattle[J]. Metabolites, 2021, 11(4): 203. DOI:10.3390/metabo11040203.
[58] SAHINGIL D, HAYALOGLU A A, SIMSEK O, et al. Changes in volatile composition, proteolysis and textural and sensory properties of white-brined cheese: effects of ripening temperature and adjunct culture[J]. Dairy Science & Technology, 2014, 94(6): 603-623. DOI:10.1007/s13594-014-0185-2.
[59] ALEWIJN M, SLIWINSKI E L, WOUTERS J T M. Production of fat-derived (flavour) compounds during the ripening of Gouda cheese[J]. International Dairy Journal, 2005, 15(6/7/8/9): 733-740. DOI:10.1016/j.idairyj.2004.09.009.




